
Our Partners
Dovetail Genomics partners with distributors, core labs, and industry partners to bring linked-read solutions to labs around the world.
Dovetail Certified Service Provider Program
Dovetail Genomics, a leader in linked-read technology, invites you to unlock the potential of the growing linked-read genomics market with our innovative partnership program. Linked-read genomics data is a key data type for epigenetic research — a prominent solution across oncology, gene regulation, and human disease research and a staple of genome assembly projects.
As the global epigenetic market is projected to grow from $2.14 billion in 2023 to over $7 billion by 2033, now is the perfect time to expand your lab’s capabilities without buying new equipment.
Why Partner with Us?
Reach More Labs: Amplify your lab’s reach with our comprehensive co-marketing support, driving local experiments to your lab by adding new technologies to your existing sequencing capabilities. Expand into multiomic approaches to epigenetic research and further de novo genome assembly and genetic variation studies.
Specialized Pricing: Gain access to exclusive, volume-based pricing that makes high-quality research more affordable.
Dedicated Support: Our specialized team is ready to assist you with rapid response times, ensuring projects run smoothly.
Offer Leading Epigenetic Solutions: Benefit from early access to our latest products and initiatives. Our customers use our assays to publish in Cell, Nature, and other high impact journals.
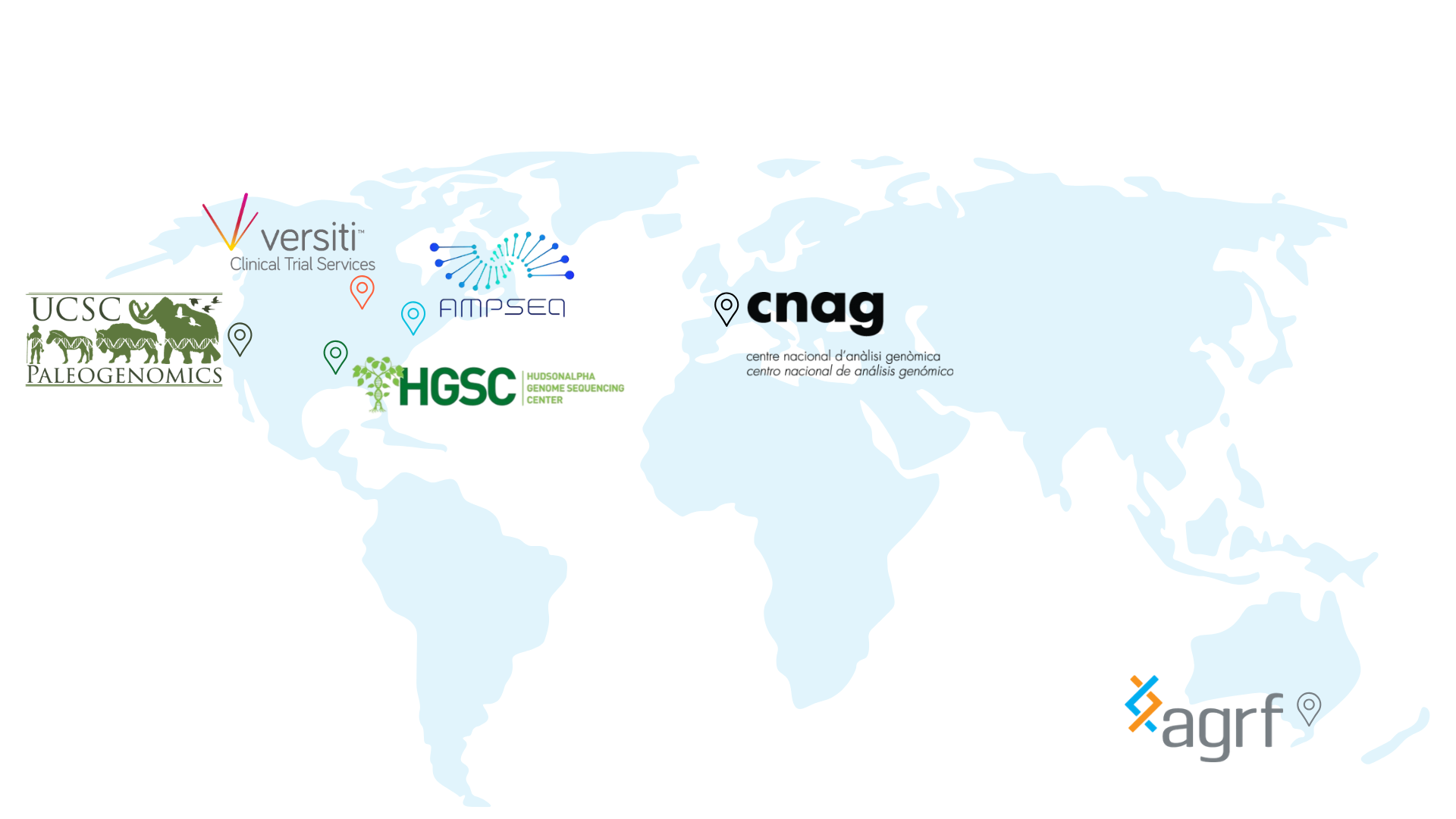
Dovetail® Certified Service Providers
The Dovetail® Certified Service Provider program underscores Dovetail Genomics’ commitment to enhancing the accessibility and quality of Hi-C data through service labs. It sets rigorous standards of proficiency and data quality that service labs must meet to achieve certification. This ensures that researchers accessing services from certified labs receive the most reliable and highest quality Hi-C data, crucial for generating high quality haplotype-resolved genome assemblies, mapping the 3D genome and discovering large and small variants across various disease states.
Australasia
Australian Genome Research Facility Ltd (AGRF)
Contact Email: advancedgenomics@agrf.org.au or CustomerCare@agrf.org.au
Website: www.agrf.org.au
Europe
Consorcio para la Explotación del Centro Nacional de Análisis Genómico (CNAG)
Contact Email: projectmanager@cnag.eu
Website: https://www.cnag.eu/
North America
AmpSeq
Contact Email: info@ampseq.com
Website: https://www.ampseq.com/
HudsonAlpha Genome Sequencing Center
Contact Email: jgrimwood@hudsonalpha.org
Website: https://gsc.hudsonalpha.org/
The Paleogenomics Lab at the University of California, Santa Cruz
Contact Email: pgl-addnap@ucsc.edu
Website: https://pgl.soe.ucsc.edu/addnap.html
Versiti Clinical Trial Services
Contact Email: clinicaltrials@versiti.org
Website: https://versiticlinicaltrials.org/
Global Distributors
-

均泰生物科技股份有限公司 Genetech Biotech Co., Ltd.
Taiwan
Address: 4F., No.168, Sinhu 2nd Rd., Neihu District,Taipei City 114-94 Taiwan
Contact Number: +886 2 2796 5108 Email:sales@gtbiotech.com.tw
Website:https://www.gtbiotech.com.tw/product/brands/13 -

Tomy Digital Biology Co., Ltd.
Japan
Address: 1-1-17 Koishikawa Bunkyo-ku Tokyo 112-0002, Japan
Contact Number: +81 3 6240 0843
Email:info_ap@digital-biology.co.jp Website:https://www.digital-biology.co.jp/allianced/products/dovetail/ -

INDNA
Republic of Korea
Address: 707ho, 636-3, Dongtan-daero, Hwaseong-si, Gyeonggi-do, 18468, Republic of Korea
Contact Number: +82-70-7514-5117
E-mail: info@indna.co.kr
Website:https://indna.co.kr/?page_id=3346 -

Bioentist Co., Ltd.
Thailand
Address: 860/9, Luangpang rd., Tubyao, Ladkrabang, Bangkok, 10250, Thailand
Contact Number:+66 2-050-9997 (Office), +66 94-440-9900 (Business)
Email:hi@bioentist.com
Website:https://bioentist.com/ -

Inqaba Biotec for Sub Saharan Africa
South Africa
Address: Menlo Park, 0102, South Africa
Contact Number: +27 12 343 5829
Email: info@inqababiotec.co.za
Website: https://inqababiotec.co.za/ -

Institute of Applied Biotechnologies a.s. (IAB)
Czech Republic, Poland, Slovakia
Address: Služeb 3056/4, 108 00 Praha-Strašnice, Czech Republic
E-mail: iabio@iabio.eu
Website: www.iabio.eu -

Biosystems Corp
Mexico, Paraguay, Uruguay
Address: Av Insurgentes Sur 1431 p 10
Colonia San José Insurgentes
Delegación Benito Juarez
Ciudad de México | MéxicoContact Number: +52 5514778719
Email: ventas@biosystemscorp.mx
Website: https://biosystemscorp.com/